

12月27 外源细胞分裂素和脱落酸响应的荔枝果皮转录组分析
影响因子:2.646
摘要
荔枝(Litchi chinensis Sonn.)是一种重要的水果作物,起源于中国,并在世界热带和亚热带地区商业化种植。荔枝红色果皮的颜色是荔枝果实市场接受度的重要品质,果皮的粉红色/红色是花青素生物合成和积累的结果,花青素-3-葡萄糖苷和花青素-3-芸香糖苷是荔枝红果皮中的主要花青素。荔枝花青素的数量和组成在不同的品种间差异很大,也很大程度上受各种环境因素的影响。因此,深入了解荔枝中花青素生物合成的调控具有重要的科学意义和经济意义。
许多研究表明,花青素生物合成很大程度上受植物激素的影响。外源脱落酸(ABA)处理促进荔枝花青素生物合成,而外源性N-(2-氯吡啶-4-基)-N’-苯基脲(CPPU)抑制荔枝花青素生物合成。但是,ABA或CPPU调控花青素生物合成的机制仍不清楚。为了进一步了解外源ABA和CPPU调控荔枝果实花青素生物合成的分子基础,华南农业大学园艺学院赵杰堂老师课题组对ABA或CPPU处理的荔枝果皮进行转录组测序,并进行了全面的分析。
材料和方法
选取生长十五年的L. chinensis cv,妃子笑。 三种处理方式:ABA处理,CPPU处理和对照处理(自来水),每组处理约15个果实簇。处理后分别取0,10和20天果皮圆盘,共取7组样本分别是:Cont-0,Cont-10,Cont-20,ABA-10,ABA-20,CPPU-10,CPPU-20,分别提取总RNA,用于RNA测序。
测序平台:北京百迈客生物科技有限公司Illumina Hiseq平台 150PE
分析平台:百迈客云平台(BMKCloud)
分析内容:1.原始数据质控;2.参考基因组比对(未发表);3.差异表达基因分析(韦恩图绘制,KEGG富集分析);4.基因功能注释(使用Blast2Go 基于Nr,Nt,COG,KO,GO数据库的最高相似性进行基因注释);5.统计分析
花青素和叶绿素含量的测定以及内源ABA分析
研究人员根据Wrolstad(1982)等人的描述进行相应的修改对果皮中总花青素含量进行定量,根据Arnon(1949)描述的方法计算总叶绿素含量,根据Jia(2011)等人所述,进行了一些修改,使用气相色谱 – 质谱(GC-MS)测定ABA含量,每个过程重复三次。
结果
1.经外源ABA和CPPU处理后,荔枝果皮的表型、色素含量和内源ABA水平的变化
处理10天后,ABA处理和对照处理的果实开始着色(图1a),尽管花青素含量没有显着差异(图1b)。然而,CPPU处理的果实只有一点点红色(图1a),并且花青素含量远远低于ABA处理和对照的果实(图1b)。处理后20天,ABA处理的果实变成均匀的红色,并且CPPU处理的果实仍然是绿色的,带有一点红色(图1a)。同时,对照果实呈现不均匀的红色(图1a),这是cv妃子笑的典型特征。如图1b所示,ABA处理20天后明显促进荔枝果皮花青素积累,而CPPU处理显着抑制荔枝果皮花青素积累。与花青素含量变化相反,果实成熟过程中叶绿素含量降低。显然,ABA处理加速叶绿素降解和CPPU处理延缓了这一过程(图1c)。ABA处理的果实与对照之间的内源ABA水平没有显着差异,而CPPU处理的果相比而言具有较低的ABA水平(图1d)。
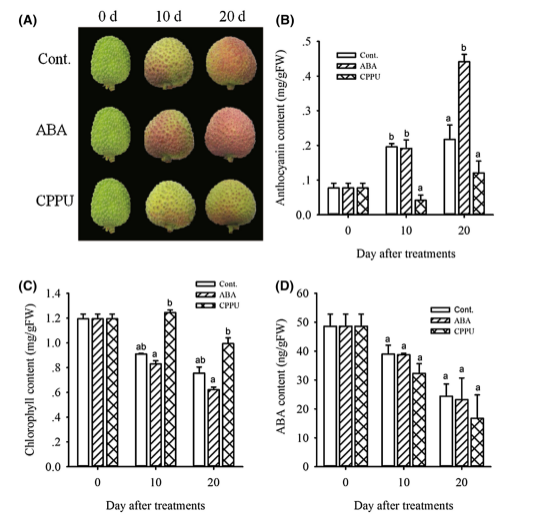
图1.各处理组果皮颜色、花青素、叶绿素及ABA含量变化
2.RNA测序以及参考基因组比对
共构建7个文库,测序过滤后每个样本数据均不低于6.6Gb,平均GC含量为45.36%(表1)。与荔枝参考基因组比对(未发表),比对率是74.7%(表1)。
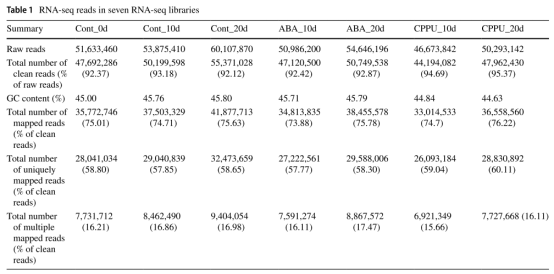
表1. RNA-Seq结果展示
3.外源ABA和CPPU处理后荔枝果皮基因表达谱的综合分析
三种处理后0天,10天和20天的荔枝果皮中发现了许多差异表达转录本(图2)。荔枝果皮色素沉着过程中,在对照组鉴定了2263个差异表达基因(DEGs)(图2a)。除此之外, 经过ABA(图2b)和CPPU(图2c)处理后的三个阶段分别鉴定了1986和2862个DEGs。
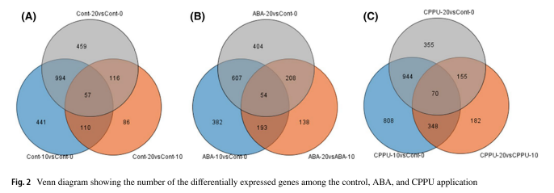
图2.三组处理中差异表达基因韦恩图
为了鉴定ABA和CPPU调控的花青素生物合成的DEGs,分别选择ABA / CPPU处理后10天和20天的DEGs与对照组进行比较。 与对照相比,在ABA和CPPU处理的果皮中分别有579和827个DEGs。 表达模式分析表明,在ABA处理10天和20天后,上调的转录本数量均比下调的转录本数量多。而 CPPU处理后20天上调的转录本数量比ABA处理的低很多。
4.外源ABA处理后的荔枝果皮表达分析
外源ABA处理组,对显著差异表达的基因进行GO分析,发现了与细胞过程,代谢过程,细胞,细胞部分,催化活性和转运蛋白活性有关的GO terms的富集。也做了DEGs的KEGG通路富集分析。 579个DEGs中的351个被富集到101个通路中,前5个通路组分别为:植物 – 病原体相互作用通路(49DEGs),植物激素信号转导(32DEGs),类黄酮生物合成(27DEGs),内质网中的蛋白质加工(27DEGs) 碳代谢(20 DEGs),淀粉和蔗糖代谢(20 DEGs)。
外源ABA处理的DEGs中,大部分与花青素合成有关的DEGs上调,如PAL,C4H,CHS,CHI,DFR,LDOX和GTs。此外, FLS和LAR等黄酮醇和原花青素合成基因上调, C3’H,COMT,F5H,CCR,CAD等木质素合成基因也上调了。 ABA处理后另一类显着差异表达的unigene参与植物激素代谢和信号传导。两个编码ABA生物合成途径中的关键酶的NCEDs unigenes显著上调,而ABA信号转导的基因在外源ABA处理后没有明显的变化。外源ABA处理也影响了与生长素信号途径有关的基因表达。例如,属于Aux / IAAs家族的两个unigenes上调。另外,编码DELLA蛋白,负调控GA信号转导的unigenes经ABA处理后上调。
5.外源CPPU处理的荔枝果皮表达分析
将CPPU处理组与对照组之间的DEG进行GO分析。 与外源ABA处理相比,差异不显着。KEGG富集分析, 897个DEGs中的493DEGs个被富集到117个通路,前六个通路组是碳代谢(54DEGs),植物 – 病原体相互作用(49DEGs),光合作用(42DEGs),氨基酸生物合成(38DEGs),苯丙素生物合成 (33DEGs),植物激素信号转导(32DEGs)。
与ABA处理不同的是,外源CPPU处理后许多上调基因参与碳代谢,氨基酸生物合成和光合作用的基因,特别是在CPPU处理后10天较明显。例如,unigene Litchi__GLEAN_10019646在外源CPPU处理10天后显着上调6.1倍,它编码捕光复合体II叶绿素a / b结合蛋白1。另外一类在外源性CPPU处理中显著差异表达的unigenes涉及叶绿素生物合成代谢。 CPPU处理后,大部分参与类黄酮和花青素生物合成的DEGs均下调,如PAL,C4H,CHS和LDOX。此外,黄酮醇和原花青素合成基因,如FLS和LAR也下调。有29个DEGs比对到植物激素信号转导途径。其中,ABA信号中的两个ABA受体PYR / PYL均被CPPU下调。另外,与生长素,GA和乙烯信号有关的基因大部分也下调。
6.ABA和CPPU处理的荔枝果皮的表达分析
受外源ABA和CPPU共同响应的DEGs共199个,其中大部分unigenes在两种处理组中显示出类似的改变。 值得注意的是,有10个unigenes在转录水平上表现出相反的模式。 其中,编码GST4蛋白的unigene(Litchi_GLEAN_10051861)被报道与荔枝中花青素的液泡转移有关。
ABA处理和CPPU处理之间的显着差异之一是叶绿素代谢。 鉴定了与叶绿素生物合成和降解有关的24个候选基因,其响应外源ABA和CPPU的表达模式如图3所示。总体而言,ABA处理对叶绿素生物合成和降解的基因表达没有显著影响。 与对照相比,CPPU处理显着提高大多数叶绿素合成基因,并下调TF SGR。
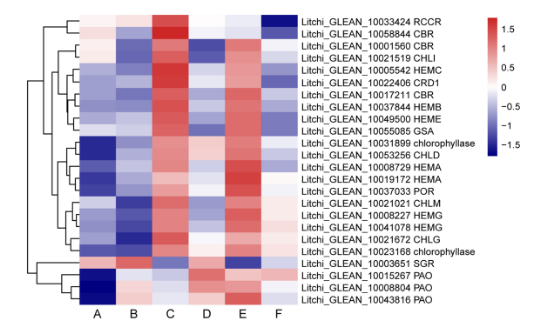
图3.两种处理后荔枝果皮中叶绿素生物合成和降解相关差异表达基因热图
经外源ABA处理和CPPU处理的一些涉及类黄酮生物合成途径的unigenes的表达有不同的改变(图4)。 该研究鉴定了不止一个荔枝黄酮类生物合成的结构基因,并且不同的基因家族成员表现出不同的表达模式。 根据研究人员以前的研究,选择黄酮生物合成相关基因作进一步分析。 如图4所示,ABA处理上调荔枝黄酮合成的结构基因,而CPPU抑制其表达,特别是PAL,CHS和F3’H。
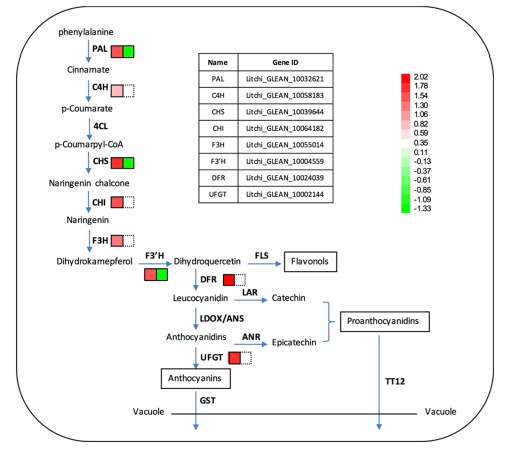
图4.两种处理后荔枝果皮中黄酮类生物合成途径相关差异表达基因概览
7.qRT-PCR验证
对9个显着差异表达的基因进行qRT-PCR实验,分别是:HEMA,CBR,DFR,CHI,UFG,chloaophyllase,RCCR和SGR。 结果显示,RT-PCR结果与RNA-seq数据一致(图5)。
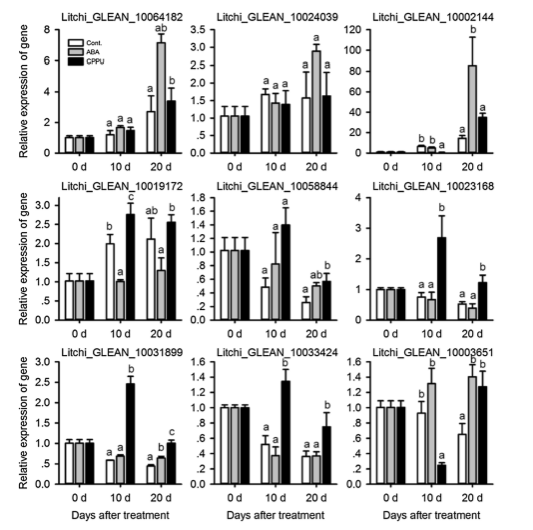
图5. qRT-PCR分析结果图
结论
在褪色阶段外源ABA处理提高了荔枝的颜色,而外源CPPU处理抑制了花青素的积累。 RNA-seq分析结果显示:两种处理组(ABA和CPPU)与对照相比,分别有579个和827个差异表达基因。外源ABA处理中,上调表达的有类黄酮和花青素合成相关基因。相反,外源CPPU处理中,诱导了与碳代谢,氨基酸生物合成和光合作用有关的基因,并下调了花青素生物合成相关基因。结果表明,ABA处理对花青素生物合成和糖代谢中涉及的基因的表达有显着作用。 另外,ABA处理对叶绿素分解代谢相关基因无显着影,CPPU处理显着增加了叶绿素合成基因的表达,并抑制了叶绿素降解基因(SGR)的表达,说明外源CPPU处理通过增加叶绿素合成基因的表达和抑制叶绿素降解基因的表达来影响叶绿素分解代谢。进一步分析显示,ABA和CPPU处理也影响其他植物激素信号传导途径(如生长素,GA和乙烯)中的基因表达,说明ABA和CPPU可能与其他激素信号途径如生长素,GA和乙烯相互作用,形成一个复杂的网络调控花青素的生物合成。
创新点
研究人员在前期的研究中为此项研究做了较好的铺垫,然而,早期的研究仅关注于外源ABA和CPPU对荔枝果皮单基因或少量基因的表达。本研究采用转录组分析方法,了解外源ABA和CPPU处理后荔枝果皮的整体分子变化。本研究为ABA和细胞分裂素影响荔枝和其他富含花青素的果树中花青素生物合成的机制探索提供了有价值的信息。
参考文献:Hu, B., Li, J., Wang, D. et al. Plant Growth Regul (2017). https://doi.org/10.1007/s10725-017-0351-7